ЗАКЛЮЧЕНИЕ
Исследование биохимических процессов при мышечной деятельности является значимым не только для спортивной биохимии, биологии, физиологии, но и для медицины, потому что профилактика переутомления, повышение возможностей организма, а также ускорение процессов восстановления являются важными аспектами сохранения и укрепления здоровья населения.
Глубокие биохимические исследования на молекулярном уровне способствуют совершенствованию методов тренировки, поиску наиболее эффективных способов повышения работоспособности, разработке путей реабилитации спортсменов, а также оценке их тренированности и рационализации питания.
При мышечной деятельности различной мощности в той или иной степени изменяются процессы обмена гормонов, которые в свою очередь регулируют развитие биохимических изменений в организме в ответ на физическую нагрузку. Важная роль принадлежит циклическим нуклеотидам как вторичных посредников гормонов и нейромедиаторов в регуляции внутриклеточного метаболизма, а также регуляции функциональной активности мышц.
Основываясь на данные литературы, мы убедились в том, что степень изменения биохимических процессов в организме зависит от типа выполняемого упражнения, его мощности и продолжительности.
Анализ специальной литературы позволил изучить биохимические изменения в организме спортсмена при мышечной работе. В первую очередь, эти изменения касаются механизмов аэробного и анаэробного энергообразования, которые зависят от типа выполняемой мышечной работы, ее мощности и продолжительности, а также от тренированности спортсмена. Биохимические изменения при мышечной деятельности наблюдаются во всех органах и тканях организма, что свидетельствует о высоком воздействии физических упражнений на организм.
По данным литературы показаны анаэробный (безкислородный) и аэробный(с участием кислорода)механизмы энергообеспечения мышечной деятельности. Анаэробный механизм в большей степени обеспечивает энергией при максимальной и субмаксимальной мощности выполнения упражнений, так как имеет достаточно высокую скорость развёртывания. Аэробный механизм выступает основным при длительной работе большой и умеренной мощности, является биохимической основой общей выносливости, поскольку его метаболическая емкость практически безгранична.
Биохимические сдвиги в организме при выполнении упражнений различной мощности определяются по содержанию продуктов метаболизма мышц в крови, моче, выдыхаемом воздухе, а также непосредственно в мышцах.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Бринзак В. П. Исследование изменений кислотно - щелочного равновесия в развитии артериальной гипоксемии при мышечной деятельности: Автореф…канд.биол.наук. - Тарту, 1979. - 18 с.
2. Виру А. А., Кырге П. К. Гормоны и спортивная работоспособность - М; Физкультура и спорт, 1983 - 159 с.
3. Волков Н. И. Адаптация энергетического метаболизма у человека к воздействию физических нагрузок при систематических занятиях спортом//Физиол.пробл.адаптации: Тез. - Тарту, 1984 - 94 с.
4. Волков Н. И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности: учебник для ИФК- Олимп.лит-ра, 2000.- 503 с.
5. Горохов А. Л. Содержание катехоламинов в крови и мышцах и их связь с биохим. изменениями в организме при мышечной деятельности//Укр.биохим.журн. - 1971- Т.43,№2 - 189 с.
6. Гусев Н. Б. Фосфорилирование миофибриллярных белков и регуляция сократительной активности//Успехи биол.химии. - 1984. - Т.25 - 27 с.
7. Калинский М. И. Состояние аденилатциклазной системы скелетных мышц при тренировке физическими нагрузками: Тр. Тартуского ун-та. - Тарту, 1982. - 49 с.
8. Калинский М. И., Кононенко В.Я. Особенности обмена катехоламинов при мышечной деятельности в тренированном организме: Материалы сов.-амер. Симп. По биохимии спорта. - Л., - 1974.- 203 с.
9. Калинский М.И.. Курский М.Д., Осипенко А.А. Биохимические механизмы адаптации при мышечной деятельности. - К.: Вища шк. Головное изд-во,1986. - 183 с.
10. Калинский М.И., Рогозкин В.А. Биохимия мышечной деятельности. - К.:Здоровья, 1989. - 144 с.
11. Курский М.Д. Транспорт кальция и роль цАМФ- зависимого фосфорилирования в его регуляции// Укр. биохим. журн. - 1981. - Т.53, №2. - 86 с.
12. Матлина Э. Ш., Кассиль Г.Н. Обмен катехоламинов при физической нагрузке у человека и животных//Успехи физиол.наук. - 1976. - Т.7,№2. - 42 с.
13. Меерсон Ф. З. Адаптация сердца к большой нагрузке и сердечная недостаточность. - М: Наука, 1975. - 263 с.
14. Меньшиков В.В. и др. Эндокринная функция поджелудочной железы при физической нагрузке//Уч. зап. Тартуского ун-та. - 1981. - Вып.562. - 146 с.
15. Панин Л. Е. Биохимические механизмы стресса. - Новосибирск: Наука, 1984. - 233 с.
16. Рогозкин В. А. О регуляции метаболизма скелетных мышц при их систематической функции//Обмен веществ и биохим. оценка тренированности спортсмена: Материалы сов. - амер. симп. - Л., 1974. - 90 с.
17. Сээне Т.П. Актомиозиновая АТФ-азная активность сердечной и скелетной мышц при физ. тренировке//Уч.зап. Тартуского ун-та. - 1980. - Вып.543. - 94 с.
18. Томсон К.Э. Влияние мышечной деятельности на тиреоидный гомеостаз организма// Уч.зап. Тартуского ун-та. - 1980. - Вып.543. -116 с.
19. Хайдарлиу С.Х. Функциональная биохимия адаптации. - Кишинев: Штиинца, 1984. - 265 с.
20. Хочачка П., Сомеро Д. Стратегия биохимической адаптации. - М: Мир, 1977. - 398 с.
21. Чернов В.Д. Обмен иода в тканях крыс при физических нагрузках//Укр. биохим. журн. - 1981. - Т.53№6. - 86 с.
22. Шмальгаузен И.И. Регуляция формообразования в индивидуальном развитии. - М: Наука. 1964. - 156 с.
23. Эллер А.К. Значение глюкокортикоидов в регуляции белкового обмена и механизм их действия в миокарде при мышечной деятельности: Автореф.дис.канд.мед. наук. - Тарту,1982. - 24 с.
24. Яковлев Н.Н. Биохимия спорта. - М: Физкультура и спорт, 1974. - 288 с.
25. Яковлев Н.Н. Влияние мышечной деятельности на белки мышц, содержание саркоплазматического ретикулума и поглощение им Са 2+ // Укр. биохим. журн. - 1978. - Т. 50, №4. - 442 с.
Мышечная система и ее функции
сокращений, общий обзор скелетной мускулатуры)
Существует два вида мускулатуры: гладкая (непроизвольная) и поперечно-полосатая (произвольная). Гладкие мышцы расположены в стенках кровеносных сосудов и некоторых внутренних органах. Они сужают или расширяют сосуды, продвигают пищу по желудочно-кишечному тракту, сокращают стенки мочевого пузыря. Поперечно-полосатые мышцы – это все скелетные мышцы, которые обеспечивают многообразные движения тела. К поперечно-полосатым мышцам относится также и сердечная мышца, автоматически обеспечивающая ритмическую работу сердца на протяжении всей жизни. Основа мышц – белки, составляющие 80–85% мышечной ткани (исключая воду). Главное свойство мышечной ткани – сократимость, она обеспечивается благодаря сократительным мышечным белкам – актину и миозину.
Мышечная ткань устроена очень сложно. Мышца имеет волокнистую структуру, каждое волокно – это мышца в миниатюре, совокупность этих волокон и образуют мышцу в целом. Мышечное волокно, в свою очередь, состоит из миофибрилл. Каждая миофибрилла разделена на чередующиеся светлые и темные участки. Темные участки – протофибриллы состоят из длинных цепочек молекул миозина, светлые образованы более тонкими белковыми нитями актина. Когда мышца находится в несокращенном (расслабленном) состоянии, нити актина и миозина лишь частично продвинуты относительно друг друга, причем каждой нити миозина противостоят, окружая ее, несколько нитей актина. Более глубокое продвижение относительно друг друга обусловливает укорочение (сокращение) миофибрилл отдельных мышечных волокон и всей мышцы в целом (рис. 2.3).
К мышце подходят и от нее отходят (принцип рефлекторной дуги) многочисленные нервные волокна (рис. 2.4). Двигательные (эфферентные) нервные волокна передают импульсы от головного и спинного мозга, приводящие мышцы в рабочее состояние; чувствительные волокна передают импульсы в обратном направлении, информируя центральную нервную, систему о деятельности мышц. Через симпатические нервные волокна осуществляется регуляция обменных процессов в мышцах, посредством чего их деятельность приспосабливается к изменившимся условиям работы, к различным мышечным нагрузкам. Каждую мышцу пронизывает разветвленная сеть капилляров, по которым поступают необходимые дли жизнедеятельности мышц вещества и выводятся продукты обмена.
Скелетная мускулатура. Скелетные мышцы входят в структуру опорно-двигательного аппарата, крепятся к костям скелета и при сокращении приводят в движение отдельные звенья скелета, рычаги. Они участвуют в удержании положения тела и его частей в пространстве, обеспечивают движения при ходьбе, беге, жевании, глотании, дыхании и т.д., вырабатывая при этом тепло. Скелетные мышцы обладают способностью возбуждаться под влиянием нервных импульсов. Возбуждение проводится до сократительных структур (миофибрилл), которые, сокращаясь, выполняют определенный двигательный акт – движение или напряжение.
Рис. 2.3. Схематическое изображение мышцы.
Мышца (Л) состоит из мышечных волокон (Б), каждое из них - из миофибрилл (В). Миофибрилла (Г) составлена из толстых и тонких миофиламентов (Д). На рисунке показан один саркомер, ограниченный с двух сторон линиями: 1 - изотропный диск, 2 - анизотропный диск, 3 - участок с меньшей анизотропностью. Поперечный сред мнофибриллы (4), дающий представление о гексагональиом распределении толстых и тонких мнофиламснтов

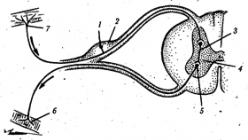
Рис. 2.4. Схема простейшей рефлекторной дуги:
1 - аффрерентный (чувствительный) нейрон, 2 - спинномозговой узел, 3 - вставочный нейрон, 4 .- серое вещество спинного мозга, 5 - эфферентный (двигательный) нейрон, 6 - двигательное нервное окончание в мышцах; 7 - чувствительное нервное окончание в коже
Напомним, что вся скелетная мускулатура состоит из поперечно-полосатых мышц. У человека их насчитывается около 600 и большинство из них - парные. Их масса составляет 35-40% общей массы тела взрослого человека. Скелетные мышцы снаружи покрыты плотной со-единительнотканной оболочкой. В каждой мышце различают активную часть (тело мышцы) и пассивную (сухожилие). Мышцы делятся на длинные, короткие и широкие.
Мышцы, действие которых направлено противоположно, называются антагонистами, однонаправленно - синергистами. Одни и те же мышцы в различных ситуациях могут выступать в том и другом качестве. У человека чаще встречаются веретенообразные и лентовидные. Веретенообразные мышцы расположены и функционируют в районе длинных костных образований конечностей, могут иметь два брюшка (двубрюшные мышцы) и несколько головок (двуглавые, трехглавые, четырехглавые мышцы). Лентовидные мышцы имеют различную ширину и обычно участвуют в корсетном образовании стенок туловища. Мышцы с перистым строением, обладая большим физиологическим поперечником за счет большого количества коротких мышечных структур, значительно сильнее тех мышц, ход волокон в которых имеет прямолинейное (продольное) расположение. Первые называют сильными мышцами, осуществляющими малоамплитудные движения, вторые - ловкими, участвующими в движениях с большой амплитудой. По функциональному назначению и направлению движений в суставах различают мышцы сгибатели и разгибатели, приводящие и отводящие, сфинктеры (сжимающие) и расширители.
Сила мышцы определяется весом груза, который она может поднять на определенную высоту (или способна удерживать при максимальном возбуждении), не изменяя своей длины. Сила мышцы зависит от суммы сил мышечных волокон, их сократительной способности; от количества мышечных волокон в мышце и количества функциональных единиц, одновременно возбуждающихся при развитии напряжения; от исходной длины мышцы (предварительно растянутая мышца развивает большую силу); от условий взаимодействия с костями скелета.
Сократительная способность мышцы характеризуется ее абсолютной силой, т.е. силой, приходящейся на 1 см 2 поперечного сечения мышечных волокон. Для расчета этого показателя силу мышцы делят на площадь ее физиологического поперечника (т.е. на сумму площадей всех мышечных волокон, составляющих мышцу). Например: в среднем у человека сила (на 1 см 2 попереченого сечения мышцы) икроножной мышцы. - 6,24; разгибателей шеи - 9,0; трехглавой мышцы плеча - 16,8кг.
Центральная нервная система регулирует силу сокращения мышцы путем изменения количества одновременно участвующих в сокращении функциональных единиц, а также частотой посылаемых к ним импульсов. Учащение импульсов ведет к возрастанию величины напряжения.
Работа мышц. В процессе мышечного сокращения потенциальная химическая энергия переходит в потенциальную механическую энергию напряжения и кинетическую энергию движения. Различают внутреннюю и внешнюю работу. Внутренняя работа связана с трением в мышечном волокне при его сокращении. Внешняя работа проявляется при перемещении собственного тела, груза, отдельных частей организма (динамическая работа) в пространстве. Она характеризуется коэффициентом полезного действия (КПД) мышечной системы, т.е. отношением производимой работы к общим энергетическим затратам (для мышц человека кпд составляет 15-20%, у физически развитых тренированных людей этот показатель несколько выше).
При статических усилиях (без перемещения) можно говорить не о работе как таковой с точки зрения физики, а о работе, которую следует оценивать энергетическими физиологическими затратами организма.
Мышца как орган. В целом мышца как орган представляет собой сложное структурное образование, которое выполняет определенные функции, состоит на 72-80% из воды и на 16-20% из плотного вещества. Мышечные волокна состоят из миофибрилл с клеточными ядрами, рибосомами, митохондриями, саркоплазматическим ретикулюмом, чувствительными нервными образованиями - проприорецепторами и другими функциональными элементами, обеспечивающими синтез белков, окислительное фосфорилирование и ресинтез аденозинтрифосфорной кислоты, транспортировку веществ внутри мышечной клетки и т.д. в процессе функционирования мышечных волокон. Важным структурно-функциональным образованием мышцы является двигательная, или нейромоторная, единица, состоящая из одного мотонейрона и иннервируемых им мышечных волокон. Различают малые, средние и большие двигательные единицы в зависимости от количества мышечных волокон, задействованных в акте сокращения.
Система соединительнотканных прослоек и оболочек связывает мышечные волокна в единую рабочую систему, обеспечивающую с помощью сухожилий передачу возникающей при мышечном сокращении тяги на кости скелета.
Вся мышца пронизана разветвленной сетью кровеносных и веточками лимфатических сосунов. Красные мышечные волокна обладают более густой сетью кровеносных сосудов, чем белые. Они имеют большой запас гликогена и липидов, характеризуются значительной тонической активностью, способностью к длительному напряжению и выполнению продолжительной динамической работы. Каждое красное волокно имеет больше, чем белое, митохондрий - генераторов и поставщиков энергии, окруженных 3-5 капиллярами, и это создает условия для более интенсивного кровоснабжения красных волокон и высокого уровня обменных процессов.
Белые мышечные волокна имеют миофибриллы, которые толще и сильнее миофибрилл красных волокон, они быстро сокращаются, но не способны к длительному напряжению. Митохондрий белого вещества имеют только один капилляр. В большинстве мышц содержатся красные и белые волокна в разных пропорциях. Различают также мышечные волокна тонические (способные к локальному возбуждению без его распространения); фазные, .способные реагировать на распространяющуюся волну возбуждения как сокращением, так и расслаблением; переходные, сочетающие оба свойства.
Мышечный насос - физиологическое понятие, связанное с мышечной функцией и ее влиянием на собственное кровоснабжение. Принципиальное его действие проявляется следующим образом: во время сокращения скелетных мышц приток артериальной крови к ним замедляется и ускоряется отток ее по венам; в период расслабления венозный отток уменьшается, а артериальный приток достигает своего максимума. Обмен веществ между кровью и тканевой жидкостью происходит через стенку капилляра.

Рис. 2.5. Схематическое изображение процессов, происходящих в
синапсе при возбуждении:
1 - синаптические пузырьки, 2 - пресинаптическая мембрана, 3 - медиатор, 4 - пост-синаптическая мембрана, 5 - синаптическая щель
Механизмы мышечного Функции мышц регулируются различными сокращения отделами центральной нервной системы (ЦНС), которые во многом определяют характер их разносторонней активности
(фазы движения, тонического напряжения и др.). Рецепторы Двигательного аппарата дают начало афферентным волокнам двигательного анализатора, которые составляют 30-50% волокон смешанных (афферентно-эфферентных) нервов, направляющихся в спинной мозг. Сокращение мышц Вызывает импульсы, которые являются источником мышечного чувства - кинестезии.
Передача возбуждения с нервного волокна на мышечное осуществляется через нервно-мышечный синапс (рис. 2.5), который состоит из двух разделенных щелью мембран - пресинаптической (нервного происхождения) и постсинаптической (мышечного происхождения). При воздействии нервного импульса выделяются кванты ацетилхолина, который приводит к возникновению электрического потенциала, способного возбудить мышечное волокно. Скорость проведения нервного импульса через синапс в тысячи раз меньше, чем в нервном волокне. Он проводит возбуждение только в направлении к мышце. В норме через нервно-мышечный синапс млекопитающих может пройти до 150 импульсов в одну секунду. При утомлении (или патологии) подвижность нервно-мышечных окончаний снижается, а характер импульсов может изменяться.
Химизм и энергетика мышечного сокращения. Сокращение и напряжение мышцы осуществляется за счет энергии, освобождающейся при химических превращениях, которые происходят при поступлении в
мышцу нервного импульса или нанесении на нее непосредственного раздражения. Химические превращения в мышце протекают как при наличии кислорода (в аэробных условиях), так и при его отсутствии (в анаэробных условиях).
Расщепление и ресинтез аденозинтрифосфорной кислоты (АТФ). Первичным источником энергии для сокращения мышцы служит расщепление АТФ (она находится в клеточной мембране, ретикулюме и миозиновых нитях) на аденозиндифосфорную кислоту (АДФ) и фосфорные кислоты. При этом из каждой грамм-молекулы АТФ освобождается 10 000 кал:
АТФ = АДФ + НзР04 + 10 000 кал.
АДФ в ходе дальнейших превращений дефосфолирируется до аде-ниловой кислоты. Распад АТФ стимулирует белковый фермент актомиозин (аденозинтрифосфотаза). В покое он не активен, активизируется при возбуждении мышечного волокна. В свою очередь АТФ воздействует на нити миозина, увеличивая их растяжимость. Активность актомиозина увеличивается под воздействием ионов Са, которые в состоянии покоя располагаются в саркоплазматическом ретикулюме.
Запасы АТФ в мышце незначительны и, чтобы поддерживать их деятельность, необходим непрерывный ресинтез АТФ. Он происходит за счет энергии, получаемой при распаде креатинфосфата (КрФ) на креатин (Кр) и фосфорную кислоту (анаэробная фаза). С помощью ферментов фосфатная группа от КрФ быстро переносится на АДФ (в течение тысячных долей секунды). При этом на каждый моль КрФ освобождается 46 кДж:
Таким образом, конечный процесс, обеспечивающий все энергетические расходы мышцы, - процесс окисления. Между тем длительная деятельность мышцы возможна лишь При достаточном поступлении к ней кислорода, так как содержание веществ, способных отдавать энергию, в анаэробных условиях постепенно падает. Кроме того, при этом накапливается молочная кислота, сдвиг реакции в кислую сторону нарушает ферментативные реакции и может привести к угнетению и дезорганизации обмена веществ и снижению работоспособности мышц. Подобные условия возникают в организме человека при работе максимальной, субмаксимальной и большой интенсивности (мощности), например при беге на короткие и средние дистанции. Из-за развившейся гипоксии (нехватки кислорода) не полностью восстанавливается АТФ, возникает так называемый кислородный долг и накапливается молочная кислота.
Аэробный ресинтез АТФ (синонимы: окислительное фосфолири-рование, тканевое дыхание) - в 20 раз эффективнее анаэробного энергообразования. Накопленная во время анаэробной деятельности и в процессе длительной работы часть молочной кислоты окисляется до углекислоты и воды (1/4-1/6 ее часть), образующаяся энергия используется на восстановление оставшихся частей молочной кислоты в глюкозу и гликоген, при этом обеспечивается ресинтез АТФ и КрФ. Энергия окислительных процессов используется также и для ресинтеза углеводов, необходимых мышце для ее непосредственной деятельности.
В целом углеводы дают наибольшее количество энергии для мышечной работы. Например, при аэробном окислении глюкозы образуются 38 молекул АТФ (для сравнения: при анаэробном распаде углевода образуется лишь 2 молекулы АТФ).
Время развертывания аэробного пути образования АТФ составляет 3-4 мин (у тренированных - до 1 мин), максимальная мощность при этом 350-450 кал/мин/кг, время поддержания максимальной мощности - десятки минут. Если в покое скорость аэробного ресинтеза АТФ невысокая, то при физических нагрузках его мощность становится максимальной и при этом аэробный путь может работать часами. Он отличается также высокой экономичностью: в ходе этого процесса идет глубокий распад исходных веществ до конечных продуктов СОг и НаО. Кроме того, аэробный путь ресинтеза АТФ отличается универсальностью в использовании субстратов: окисляются все органические вещества организма (аминокислоты, белки, углеводы, жирные кислоты, кетоновые тела и др.).
Однако аэробный способ ресинтеза АТФ имеет и недостатки: 1) он требует потребления кислорода, доставка которого в мышечную ткань обеспечивается дыхательной и сердечно-сосудистой системами, что, естественно, связано с их напряжением; 2) любые факторы, влияющие на состояние и свойство мембран митохондрий, нарушают образование АТФ; 3) развертывание аэробного образования АТФ продолжительно во времени и невелико по мощности.
Мышечная деятельность, осуществляемая в большинстве видов спорта, не может полностью быть обеспечена аэробным процессом ре-синтеза АТФ, и организм вынужден дополнительно включать анаэробные способы образования АТФ, имеющие более короткое время развертывания и большую максимальную мощность процесса (т.е. наибольшее количество АТФ," образуемое в единицу времени) - 1 моль АТФ соответствует 7,3 кал, или 40 Дж (1 кал == 4,19 Дж).
Возвращаясь к анаэробным процессам энергообразования, следует уточнить, что они протекают по меньшей мере в виде двух типов реакций: 1. Креатинфосфокиназная - когда осуществляется расщепление КрФ, фосфорные группировки с которого переносятся на АДФ, ресинтезируя при этом АТФ. Но запасы креатинфосфата в мышцах невелики и это обусловливает быстрое (в течение 2-4 с) угасание этого типа реакции. 2. Гликолитическая (гликолиз) - развивается медленнее, в течение 2-3 мин интенсивной работы. Гликолиз начинается с фосфолирирования запасов гликогена мышц и поступающей с кровью глюкозы. Энергии этого процесса хватает на несколько минут напряженной работы. На этом этапе завершается первая стадия фосфолирирования гликогена и происходит подготовка к окислительному процессу. Затем наступает вторая стадия гликолитической реакции - дегидрогенирование и третья - восстановление АДФ в АТФ. Гликолитическая реакция заканчивается образованием двух молекул молочной кислоты, после чего разворачиваются дыхательные процессы (к 3-5 мин работы), когда начинает окисляться молочная кислота (лак-тат), образованная в процессе анаэробных реакций.
Биохимическими показателями оценки креатинфосфатного анаэробного пути ресинтеза АТФ является креатининовый коэффициент и алактатный (без молочной кислоты) кислородный долг. Креатининовый коэффициент - это выделение креатинина с мочой за сутки в расчете на 1 кг массы тела. У мужчин выделение креатинина колеблется в пределах 18-32 мг/сут х кг, а у женщин - 10-25 мг/сут х кг. Между содержанием креатинфосфата и образованием у него креатинина существует прямолинейная зависимость. Следовательно, с помощью креатининового коэффициента можно оценить потенциальные возможности этого пути ресинтеза АТФ.
Биохимические сдвиги в организме, обусловленные накоплением молочной кислоты в результате гликолиза. Если в покое до начала мы шечной деятельности концентрация лактата в крови составляет 1- 2 ммоль/л, то после интенсивных, непродолжительных нагрузок в течение 2-3 мин эта величина может достигать 18-20 ммоль/л. Другим показателем, отражающим накопление в крови молочной кислоты, служит показатель крови (рН): в покое 7,36, после нагрузки снижение до 7,0 и более. Накопление лактата в крови определяет и ее щелочной резерв - щелочные компоненты всех буферных систем крови.
Окончание интенсивной мышечной деятельности сопровождается снижением потребления кислорода - вначале резко, затем более плавно. В связи с этим выделяют два компонента кислородного долга: быстрый (алактатный) и медленный (лактатный). Лактатный - это то количество кислорода, которое используется после окончания работы для устранения молочной кислоты: меньшая часть окисляется до J-bO и СОа, большая часть превращается в гликоген. На это превращение тратится значительное количество АТФ, которая образуется аэробным путем за счет кислорода, составляющего лактатный долг. Метаболизм лактата осуществляется в клетках печени и миокарда.
Количество кислорода, необходимое для полного обеспечения выполняемой работы, называют кислородным запросом. Например, в беге на 400 м кислородный запрос, равен приблизительно 27 л. Время про-бегания дистанции на уровне мирового рекорда составляет около 40 с. Исследования показали, что за это время спортсмен поглощает 3-4 л 02. Следовательно, 24 л - это общий кислородный долг (около 90% кислородного запроса), который ликвидируется после забега.
В беге на 100 м кислородный долг может доходить до 96% запроса. В беге на 800 м доля анаэробных реакций несколько снижается - до 77%, в беге на 10 000 м - до 10%, т.е. преобладающая часть энергии поставляется за счет дыхательных (аэробных) реакций.
Механизм мышечного расслабления. Как только в мышечное волокно перестают поступать нервные импульсы, ионы Са^ под действием так называемого кальциевого насоса за счет энергии АТФ уходят в цистерны саркоплазматического ретикулюма и их концентрация в саркоплазме понижается до исходного уровня. Это вызывает изменения конформации тропонина, который, фиксируя тропомиозин в определенном участке актиновых нитей, делает невозможным образование поперечных мостиков между толстыми и тонкими нитями. За счет упругих сил, возникающих при мышечном сокращении в коллагеновых нитях, окружающих мышечное волокно, оно при расслаблении возвращается в исходное состояние. Таким образом, процесс мышечного расслабления, или релаксации, так же, как и процесс мышечного сокращения, осуществляется с использованием энергии гидролиза АТФ.
В ходе мышечной деятельности в мышцах поочередно происходят процессы сокращения и расслабления и, следовательно, скоростно-силовые качества мышц в равной мере зависят от скорости мышечного сокращения и от способности мышц к релаксации.
Краткая характеристика гладких мышечных волокон. В гладких мышечных волокнах отсутствуют миофибриллы. Тонкие нити (актиновые) соединены с сарколеммой, толстые (миозиновые) находятся внутри мышечных клеток. В гладких мышечных волокнах отсутствуют также цистерны с ионами Са. Под действием нервного импульса ионы Са медленно поступают в саркоплазму из внеклеточной жидкости и также медленно уходят после того, как прекращают поступать нервные импульсы. Поэтому гладкие мышечные волокна медленно сокращаются и медленно расслабляются.
Общий обзор скелетных мышц человека. Мышцы туловища (рис. 2.6 и 2.7) включают мышцы грудной клетки, спины и живота. Мышцы грудной клетки участвуют в движениях верхних конечностей, а также обеспечивают произвольные и непроизвольные дыхательные движения. Дыхательные мышцы грудной клетки называются наружными и внутренними межреберными мышцами. К дыхательным мышцам относится также и диафрагма. Мышцы спины состоят из поверхностных и глубоких мышц. Поверхностные обеспечивают некоторые движения верхних конечностей, головы и шеи. Глубокие («выпрямители туловища») прикрепляются к остистым отросткам позвонков и тянутся вдоль позвоночника. Мышцы спины участвуют в поддержании вертикального положения тела, при сильном напряжении (сокращении) вызывают прогибание туловища назад. Брюшные мышцы поддерживают давление внутри брюшной полости (брюшной пресс), участвуют в некоторых движениях тела (сгибание туловища вперед, наклоны и повороты в стороны), в процессе дыхания.
Мышцы головы и шеи - мимические, жевательные и приводящие в движение голову и шею. Мимические мышцы прикрепляются одним своим концом к кости, другим - к коже лица, некоторые могут начинаться и оканчиваться в коже. Мимические мышцы обеспечивают движения кожи лица, отражают различные психические состояния человека, сопутствуют речи и имеют значение в общении. Жевательные мышцы при сокращении вызывают движение нижней челюсти вперед и в стороны. Мышцы шеи участвуют в движениях головы. Задняя группа мышц, в том числе и мышцы затылка, при тоническом (от слова «тонус») сокращении удерживает голову в вертикальном положении.

Рис. 2.6. Мышцы передней половины тела (по Сыльвановичу):
1 - височная мышца, 2 - жевательная мышца, 3 - грудино-ключично-сосцевидная мышца, 4 - большая грудная мышца, 5 - средняя лестничная мышца, б - наружная косая мышца живота, 7 - медиальная широкая мышца бедра, 8 - латеральная широкая мышца бедра, 9 - прямая мышца бедра, 10 - портняжная мышца, 11 - нежная мышца, 12 - внутренняя косая мышца живота, 13 - прямая мышца живота, 14 - двуглавая Мышца плеча, 15 ~ наружные межреберные мышцы, 16 - круговая мышца рта, 17 - круговая мышца глаза, 18 - лобная мышца
Мышцы верхних конечностей обеспечивают движения плечевого пояса, плеча, предплечья и приводят в движение кисть и пальцы. Главными мышцами-антагонистами являются двуглавая (сгибатель) и трехглавая (разгибатель) мышцы плеча. Движения верхней конечности и прежде всего кисти чрезвычайно многообразны. Это связано с тем, что рука служит человеку органом труда.

Рис. 2.7. Мышцы задней половины тела (по Сыльвановичу):
1 - ромбовидная мышца, 2 - выпрямитель туловища, 3 - глубокие мышцы ягодичной мышцы, 4 - двуглавая мышца бедра, 5 - икроножная мышца, 6 - ахиллово сухожилие, 7 - большая ягодичная мышца, 8 - широчайшая мышца скипы, 9 - дельтовидная мышца, 10 - трапециевидная мышца
Мышцы нижних конечностей обеспечивают движения бедра, голени и стопы. Мышцы бедра играют важную роль в поддержании вертикального положения тела, но у человека они развиты сильнее, чем у других позвоночных. Мышцы, осуществляющие движения голени, расположены на бедре (например, четырехглавая мышца, функцией которой является разгибание голени в коленном суставе; антагонист этой мышцы - двуглавая мышца бедра). Стопа и пальцы ног приводятся в движение мышцами, расположенными на голени и стопе. Сгибание пальцев стопы осуществляется при сокращении мышц, расположенных на подошве, а разгибание - мышцами передней поверхности голени и стопы. Многие мышцы бедра, голени и стопы принимают участие в поддержании тела человека в вертикальном положении.
Как происходит адаптация организма спортсмена к интенсивной мышечной деятельности?
Глубокие функциональные изменения организма, возникшие в процессе адаптации его к повышенной мышечной деятельности, изучает физиология спорта. Однако в их основе лежат биохимические изменения обмена веществ тканей и органов и, в конечном итоге – организма в целом. Однако мы рассмотрим в самом общем виде основные изменения, возникающие под влиянием тренировки только в мышцах.
В основе биохимической перестройки мышц под влиянием тренировки лежит взаимозависимость процессов расходования и восстановления функциональных и энергетических резервов мышц. Как вам уже понятно из предыдущего, во время мышечной деятельности происходит интенсивное расщепление АТФ и соответственно интенсивно расходуются и другие вещества. В мышцах – это креатинфосфат, гликоген, липиды, в печени происходит расщепление гликогена с образованием сахара, который с кровью переносится к работающим мышцам, сердцу, головному мозгу; усиленно расщепляются жиры и окисляются жирные кислоты. Одновременно в организме накапливаются продукты обмена веществ – фосфорная и молочная кислоты, кетоновые тела, углекислый газ. Частично они теряются организмом, а частично используются вновь, вовлекаясь в обмен веществ. Мышечная деятельность сопровождается увеличением активности многих ферментов и благодаря этому начинается синтез израсходованных веществ. Ресинтез АТФ, креатинфосфата и гликогена возможен уже и во время работы, однако наряду с этим идет и интенсивное расщепление этих веществ. Поэтому содержание их в мышцах во время работы никогда не доходит до исходного.
В периоде отдыха, когда интенсивное расщепление источников энергии прекращается, процессы ресинтеза приобретают явный перевес и происходит не только восстановление затраченного (компенсация), но и сверхвосстановление (суперкомпенсация), превышающее исходный уровень. Эта закономерность получила название «закон суперкомпенсации».
Сущность явления суперкомпенсации.
В биохимии спорта изучены закономерности этого процесса. Установлено, например, что в случае, если происходит интенсивное расходование вещества в мышцах, в печени и др. органах, тем быстрее идет ресинтез и тем значительнее выражено явление сверхвосстановления. Например, после кратковременной интенсивной работы повышение уровня гликогена в мышцах сверх исходного наступает уже после 1 часа отдыха, а через 12 часов возвращается к исходному, дорабочему уровню. После работы большой длительности суперкомпенсация наступает только через 12 часов, но зато повышенный уровень гликогена в мышцах сохраняется более трех суток. Это возможно только благодаря высокой активности ферментов и их усиленного синтеза.
Таким образом, одной из биохимических основ изменения организма под влиянием тренировки является повышение активности ферментных систем и суперкомпенсация источников энергии, затрачиваемых во время работы. Почему закономерности суперкомпенсации очень важно учитывать в практике спортивной тренировки?
Знание закономерностей суперкомпенсации позволяет научно обосновать интенсивность нагрузок и интервалы отдыха во время обычных физических упражнений и при спортивных тренировках.
Поскольку суперкомпенсация сохраняется некоторое время по окончании работы, последующая работа может совершаться в более выгодных биохимических условиях, и, в свою очередь, приводить к дальнейшему повышению функционального уровня (рис…). Если же последующая работа совершается в условиях неполного восстановления, то это приводит к понижению функционального уровня (рис…).
Под влиянием тренировки в организме происходит активное приспособление, но не к работе «вообще», а к конкретным видам ее. При изучении различных видов спортивной деятельности был установлен принцип специфичности биохимической адаптации и установлены биохимические основы качеств двигательной деятельности – быстроты, силы, выносливости. А это означает научно обоснованные рекомендации для целенаправленной системы тренировки.
Приведем только один пример. Вспомните, как после интенсивной скоростной нагрузки (бег) наступает усиление дыхание («одышка»). С чем это связано? Во время совершения работы (бег) из-за недостаточности кислорода в крови скопились недоокисленные продукты (молочная кислота и др.), а также углекислый газ, что приводит к изменению степени кислотности крови. Соответственно это вызывает возбуждение дыхательного центра в продолговатом мозгу и усиление дыхания. В результате интенсивного окисления нормализуется кислотность крови. А это возможно только при высокой активности ферментов аэробного окисления. Следовательно, по окончании интенсивной работы в периоде отдыха активно функционируют ферменты аэробного окисления. В то же время от активности аэробного окисления напрямую зависит выносливость спортсменов, выполняющих работу большой длительности. На этом основании именно биохимики рекомендовали включать в тренировки многих видов спорта кратковременные нагрузки высокой интенсивности, что в настоящее время общепринято.
Какова биохимическая характеристика тренированного организма?
В мышцах тренированного организма:
Увеличивается содержание миозина, число свободных HS- групп в нем, т.е. способность мышц к расщеплению АТФ;
Увеличиваются запасы источников энергии, необходимых для ресинтеза АТФ (содержание креатинфосфата, гликогена, липидов и др.)
Значительно повышается активность ферментов, катализирующих, как анаэробный, так и аэробный окислительные процессы;
Возрастает содержание в мышцах миоглобина, что создает в мышцах резерв кислорода.
Увеличивается содержание белков мышечной стромы, обеспечивающей механику расслабления мышцы. Наблюдения на спортсменах показывают, что способность к расслаблению мышц под влиянием тренировки возрастают.
Адаптация к одному фактору повышает устойчивость к другим факторам (например, к стрессам и пр.);
Тренировка современного спортсмена требует высокой интенсивности физических нагрузок и большого объема их, что может оказывать одностороннее влияние на организм. Поэтому она требует постоянного контроля врачей, специалистов по спортивной медицине, опирающейся на биохимию и физиологию спорта.
И занятия физкультурой, как и спортивная деятельность, позволяют развить резервные возможности организма человека и обеспечить ему полноценное здоровье, высокую работоспособность и долголетие. Физическое здоровье составляет неотъемлемую часть гармоничного развития личности человека, формирует характер, устойчивость психических процессов, волевые качества и др.
Основоположником научной системы физического воспитания и врачебно-педагогического контроля в физической культуре является замечательный отечественный ученый, выдающийся педагог, анатом и врач Петр Францевич Лесгафт. В основе его теории лежит принцип единства физического и умственного, нравственного и эстетического развития человека. Теорию физического воспитания он рассматривал как «филиальную ветвь биологической науки».
Огромная роль в системе биологических наук, изучающих основы занятий в области физической культуры и спорта, принадлежит биохимии.
Уже в 40-х годах ушедшего века в лаборатории ленинградского ученого Николая Николаевича Яковлева были начаты целенаправленные научные исследования в области биохимии спорта. Они позволили выяснить сущность и специфические особенности адаптации организма к различным видам мышечной деятельности, обосновать принципы спортивной тренировки, факторы, влияющие на работоспособность спортсмена, на состояния утомления, перетренировки и мн. др. В дальнейшем развитие биохимии спорта составило основу подготовки космонавтов к космическим полетам.
Какие вопросы решает биохимия спорта?
Биохимия спорта является основой физиологии спорта и спортивной медицины. В биохимических исследованиях работающих мышц установлены:
Закономерности биохимических изменений как активной адаптации к повышенной мышечной деятельности;
Обоснования принципов спортивной тренировки (повторности, регулярности, соотношение работы и отдыха и др.)
Биохимическая характеристика качеств двигательной деятельности (быстроты, силы, выносливости)
Способы ускорения восстановления организма спортсмена и мн. др.
Вопросы и задания.
Почему скоростные нагрузки действуют на организм более разносторонне?
Попробуйте дать физиологическое и биохимическое обоснование высказыванию Аристотеля «Ни что так не истощает и не разрушает человека, как продолжительное физическое бездействие». Почему оно так актуально для современного человека?
В учебнике изложены основы общей биохимии и биохимии мышечной деятельности организма человека, описаны химическое строение и процессы метаболизма наиболее важных веществ организма, раскрыта их роль в обеспечении мышечной деятельности. Рассмотрены биохимические аспекты процессов мышечного сокращения и механизмов энергообразования в мышцах, закономерности развития двигательных качеств, процессов утомления, восстановления, адаптации, а также рационального питания и диагностики функционального состояния спортсменов. Для студентов и преподавателей высших и средних учебных заведений физического воспитания и спорта, специалистов по физической реабилитации и рекреации.
Информация о книге:
Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности. 2000. - 503 с.
Часть первая. Биохимические основы жизнедеятельности организма человека
Глава 1. Введение в биохимию
1. Предмет и методы исследования биохимии
2. История развития биохимии и становление биохимии спорта
3. Химическое строение организма человека
4. Превращение макромолекул
Контрольные вопросы
Глава 2. Обмен веществ в организме
1. Обмен веществ - необходимое условие существования живого организма
2. Катаболические и анаболические реакции - две стороны обмена веществ
3. Виды обмена веществ
4. Этапы распада питательных веществ и извлечения энергии в клетках
5. Клеточные структуры и их роль в обмене веществ
6. Регуляция обмена веществ
Контрольные вопросы
Глава 3. Обмен энергии в организме
1. Источники энергии
2. АТФ - универсальный источник энергии в организме
3. Биологическое окисление - основной путь энергообразования в клетках организма
4. Митохондрии - "энергетические станции" клетки
5. Цикл лимонной кислоты - центральный путь аэробного окисления питательных веществ
6. Дыхательная цепь
7. Окислительное фосфорилирование - основной механизм синтеза АТФ
8. Регуляция обмена АТФ
Контрольные вопросы
Глава 4. Обмен воды и минеральных веществ
1. Вода и её роль в организме
2. Водный баланс и его изменение при мышечной деятельности
3. Минеральные вещества и их роль в организме
4. Обмен минеральных веществ при мышечной деятельности
Контрольные вопросы
Глава 5. Кислотно-основное состояние организма
1. Механизмы транспорта веществ
2. Кислотно-основное состояние внутренней среды организма
3. Буферные системы и их роль в поддержании постоянства рН среды
Контрольные вопросы
Глава 6. Ферменты - биологические катализаторы
1. Общее представление о ферментах
2. Строение ферментов и коферментов
3. Множественные формы ферментов
4. Свойства ферментов
5. Механизм действия ферментов
6. Факторы, влияющие на действие ферментов
7. Классификация ферментов
Контрольные вопросы
Глава 7. Витамины
1. Общее представление о витаминах
2. Классификация витаминов
3. Характеристика жирорастворимых витаминов
4. Характеристика водорастворимых витаминов
5. Витаминоподобные вещества
Контрольные вопросы
Глава 8. Гормоны - регуляторы обмена веществ
1. Общее представление о гормонах
2. Свойства гормонов
3. Химическая природа гормонов
4. Регуляция биосинтеза гормонов
5. Механизм действия гормонов
6. Биологическая роль гормонов
7. Роль гормонов в мышечной деятельности
Контрольные вопросы
Глава 9. Биохимия углеводов
1. Химический состав и биологическая роль углеводов
2. Характеристика классов углеводов
3. Обмен углеводов в организме человека
4. Расщепление углеводов в процессе пищеварения и их всасывание в кровь
5. Уровень глюкозы в крови и его регуляция
6. Внутриклеточный обмен углеводов
7. Обмен углеводов при мышечной деятельности
Контрольные вопросы
Глава 10. Биохимия липидов
1. Химический состав и биологическая роль липидов
2. Характеристика классов липидов
3. Обмен жиров в организме
4. Расщепление жиров в процессе пищеварения и их всасывание
5. Внутриклеточный обмен жиров
6. Регуляция обмена липидов
7. Нарушение обмена липидов
8. Обмен жиров при мышечной деятельности
Контрольные вопросы
Глава 11. Биохимия нуклеиновых кислот
1. Химическое строение нуклеиновых кислот
2. Структура, свойства и биологическая роль ДНК
3. Структура, свойства и биологическая роль РНК
4. Обмен нуклеиновых кислот
Контрольные вопросы
Глава 12. Биохимия белков
1. Химический состав и биологическая роль белков
2. Аминокислоты
3. Структурная организация белков
4. Свойства белков
5. Характеристика отдельных белков, участвующих в обеспечении мышечной работы
6. Свободные пептиды и их роль в организме
7. Обмен белков в организме
8. Расщепление белков в процессе пищеварения и всасывание аминокислот
9. Биосинтез белка и его регуляция
10. Внутритканевой распад белков
11. Внутриклеточное превращение аминокислот и синтез мочевины
12. Обмен белков при мышечной деятельности
Контрольные вопросы
Глава 13. Интеграция и регуляция обмена веществ - биохимическая основа процессов адаптации
1. Взаимопревращение углеводов, жиров и белков
2. Регуляторные системы обмена веществ и их роль в адаптации организма к физическим нагрузкам
3. Роль отдельных тканей в интеграции промежуточного обмена веществ
Контрольные вопросы
Часть вторая. Биохимия спорта
Глава 14. Биохимия мышц и мышечного сокращения
1. Типы мышц и мышечных волокон
2. Структурная организация мышечных волокон
3. Химический состав мышечной ткани
4. Структурные и биохимические изменения в мышцах при сокращении и расслаблении
5. Молекулярный механизм мышечного сокращения
Контрольные вопросы
Глава 15. Биоэнергетика мышечной деятельности
1. Общая характеристика механизмов энергообразования
2. Креатинфосфокиназный механизм ресинтеза АТФ
3. Гликолитеческий механизм ресинтеза АТФ
4. Миокиназный механизм ресинтеза АТФ
5. Аэробный механизм ресинтеза АТФ
6. Подключение энергетических систем при различных физических нагрузках и их адаптация в процессе тренировки
Контрольные вопросы
Глава 16. Биохимеческие изменения в организме при выполнении упражнений различной мощности и продолжительности
1. Общая направленность изменения биохимических процессов при мышечной деятельности
2. Транспорт кислорода к работающим мышцам и его потребление при мышечной деятельности
3. Биохимические изменения в отдельных органах и тканях при мышечной работе
4. Классификация физических упражнений по характеру биохимических изменений при мышечной работе
Контрольные вопросы
Глава 17. Биохимические факторы утомления
1. Биохимические факторы утомления при выполнении кратковременных упражнений максимальной и субмаксимальной мощности
2. Биохимические факторы утомления при выполнении длительных упражнений большой и умеренной мощности
Контрольные вопросы
Глава 18. Биохимическая характеристика процессов восстановления при мышечной деятельности
1. Динамика биохимических процессов восстановления после мышечной работы
2. Последовательность восстановления энергетических запасов после мышечной работы
3. Устранение продуктов распада в период отдыха после мышечной работы
4. Использование особенностей протекания восстановительных процессов при построении спортивной тренировки
Контрольные вопросы
Глава 19. Биохимические факторы спортивной работоспособности
1. Факторы, лимитирующие физическую работоспособность человека
2. Показатели аэробной и анаэробной работоспособности спортсмена
3. Влияние тренировки на работоспособность спортсменов
4. Возраст и спортивная работоспособность
Контрольные вопросы
Глава 20. Биохимические основы скоростно-силовых качеств спортсмена и методы их развития
1. Биохимическая характеристика скоростно-силовых качеств
2. Биохимические основы методов скоростно-силовой подготовки спортсменов
Контрольные вопросы
Глава 21. Биохимические основы выносливости спортсменов
1. Биохимические факторы выносливости
2. Методы тренировки, способствующие развитию выносливости
Контрольные вопросы
Глава 22. Закономерности биохимической адаптации в процессе спортивной тренировки
1. Физические нагрузки, адаптация и тренировочный эффект
2. Закономерности развития биохимической адаптации и принципы тренировки
3. Специфичность адаптационных изменений в организме при тренировке
4. Обратимость адаптационных изменений при тренировке
5. Последовательность адаптационных изменений при тренировки
6. Взаимодействие тренировочных эффектов в процессе тренировки
7. Цикличность развития адаптации в процессе тренировки
Контрольные вопросы
Глава 23. Биохимические основы рационального питания спортсменов
1. Принципы рационального питания спортсменов
2. Энергопотребление организма и его зависимость от выполняемой работы
3. Сбалансированность питательных веществ в рационе спортсмена
4. Роль отдельных химических компонентов пищи в обеспечении мышечной деятельности
5. Пищевые добавки и регулирование массы тела
Контрольные вопросы
Глава 24. Биохимический контроль в спорте
1. Задачи, виды и организация биохимического контроля
2. Объекты исследования и основные биохимические показатели
3. Основные биохимические показатели состава крови и мочи, их изменение при мышечной деятельности
4. Биохимический контроль развития систем энергообеспечения организма при мышечной деятельности
5. Биохимический контроль за уровнем тренированности, утомления и восстановления организма спортсмена
6. Контроль за применением допинга в спорте
Контрольные вопросы
Словарь терминов
Единицы измерения
Литература
Дополнительно о книге: формат: pdf, размер файла: 37,13 Мб.